Typical flowers have sepals and petals surrounding male (stamen) and female (pistil or gynoecium) reproductive organs. In different plants, the forms and sizes of flower organs support the species’ mating system and mode of pollination. For example, self-pollinating plants typically have perfect (hermaphrodite) flowers whose mature stamens and pistils are of similar length to facilitate self-pollination. Other species that normally outcross may have divergent male and female organ sizes and spatial arrangement, or even separate flowers, to enable cross-pollination by animals or wind.
Flower organs must mature at an appropriate seasonal and diurnal time, for example when pollinators are active. In many species, as flowers open, petals, stamens, and/or style grow exceptionally quickly and transiently, sometimes doubling in size within a few hours. Such rapid growth creates mechanical and regulatory challenges. Due to their surrounding cell walls, plant cells cannot reorganize to accommodate growth of nearby cells, yet organs grow coherently without buckling or breaking. Growth of multiple cell layers within an organ must therefore be coupled through chemical and/or mechanical signals. Moreover, growth and differentiation of different flower organs just before flower opening must be coordinated to achieve compatible geometry and timely anther dehiscence (pollen release).
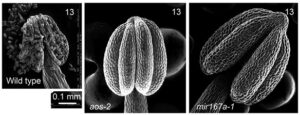

In the self-pollinating model plant Arabidopsis, two Auxin Response Factors, ARF6 and ARF8, promote flower maturation. These factors mediate transcriptional responses to the hormone auxin. The regulatory network that coordinates flower maturation also includes a second hormone, jasmonate, that is required for male fertility. Loss of ARF6 and ARF8 leads to arrest of unopened flower buds (resembling broccoli florets). Conversely, ectopic ARF6 and ARF8 activity, arising from loss of microRNA regulation, causes excess anther growth leading to failure to release pollen. Productive flower maturation thus requires both temporal and spatial regulation of auxin response. Alterations in these genes also cause parthenocarpic fruit growth (without fertilization or seed set), a potentially valuable agronomic trait.
We are currently using confocal microscopy, transcriptomic, and gene editing approaches to understand how flower organs grow rapidly without breaking or buckling, how this growth is coupled to jasmonate production, and how environmental conditions may influence these processes.
Selected relevant publications:
Israeli, A., Schubert, R., Man, N., Teboul, N., Serrani Yarce, J. C., Rosowski, E. E., Wu, M.-F., Levy, M., Efroni, I., Ljung, K., Hause, B., Reed, J. W., Ori, N. 2023. Modulating auxin response stabilizes tomato fruit set. Plant Physiology 192:2336-2355. https://doi.org/10.1093/plphys/kiad205.
Zheng, L., Nagpal, P., Villarino, G., Trinidad, B., Bird, L., Huang, Y., Reed, J.W. 2019. miR167 limits anther growth to potentiate anther dehiscence. Development 146, dev174375. doi:10.1242/dev.174375. https://dev.biologists.org/content/146/14/dev174375
Reed J.W., Wu M.-F., Reeves P.H., Hodgens C., Yadav V., Hayes S., Pierik R. 2018. Three Auxin Response Factors promote hypocotyl elongation. Plant Physiology 178(2):864-875. doi: 10.1104/pp.18.00718. http://www.plantphysiol.org/content/178/2/864.long
Reeves, P.H., Ellis, C.M., Ploense, S.E., Wu, M.-F., Yadav, V., Tholl, D., Chételat, A., Haupt, I., Kennerley, B.J., Hodgens, C., Farmer, E.E., Nagpal, P., Reed, J.W. 2012. A regulatory network for coordinated flower maturation. PLoS Genetics 8(2): e1002506. doi:10.1371/journal.pgen.1002506. https://journals.plos.org/plosgenetics/article?id=10.1371/journal.pgen.1002506
Wu, M.-F., Tian, Q., and Reed, J. W. 2006. Arabidopsis miR167 controls patterns of ARF6 and ARF8 expression and regulates both female and male reproduction. Development 133: 4211-4218. https://doi: 10.1242/dev.02602. https://journals.biologists.com/dev/article/133/21/4211/43323/Arabidopsis-microRNA167-controls-patterns-of-ARF6
Nagpal, P., Ellis, C. M., Weber, H., Ploense, S. E., Barkawi, L. S., Guilfoyle, T. J., Hagen, G., Alonso, J. M., Cohen, J. D., Farmer, E. E., Ecker, J. R., and Reed, J. W. 2005. Auxin Response Factors ARF6 and ARF8 promote jasmonic acid production and flower maturation. Development 132: 4107-4118. https://dev.biologists.org/content/132/18/4107.long